JBRA Assist. Reprod. 2012;16(5):290-300
CASE REPORT
doi: 10.5935/1518-0557.2012.16.5.07
PGD by aCGH and QF-PCR in a couple with recurring aneuploidies
1andressagmondadori@gmail.com
2mariaducatelli@yahoo.com.ar
3juka_mi@hotmail.com
4aligalo@yahoo.com.ar
5fabiancoco@fecunditas.com.ar
6Corresponding Author
7Larrea 790, CABA, Postal Code 1030, Argentina
ABSTRACT
This case report describes a PGS study perfomed on a blastocyst stage of a 44 year old female and 46 year old male couple with a history of recurrent aneuploid abortions. The main purpose of this report is to determine, by using STRs and QF-PCR, the parental origin of the aneuploidies found on the embryos aCGH karyotype. Only one blastocyst of the eight biopsed resulted cytogenetically normal and it was transferred in a deferred cycle. The patient achieved the pregnancy and delivered at term a normal male child. In 30 of the 38 aneuploidies identified we could determine the parental origin: eight maternal trisomies, eight paternal trisomies and 14 maternal monosomies. Five of the eight maternal trisomies showed an allelic pattern malsegregation 1:1:1 and three showed the pattern 2:1. The same segregation patterns were observed for the paternal origin trisomies. We conclude that the aneuploidies were contributed by both members of the couple in equal proportions. This outcome compels us to reflect that caution is required when we are offering oocyte donation in a women older than 40 years who has a partner of the similar age.
Keywords: PGS, trophectoderm biopsy, advanced paternal age, recurrent abortions, aCGH.
RESUMO
Trata-se de relato de PGS study realizado em blastocistos de mulher de 44 anos com marido de 46 anos, casal com história de abortamentos recorrentes. O principal objetivo deste é determinar, utilizando STRs e QF-PCR ,a origem parental das aneuploidias encontradas no aCGH. Apenas um blastocisto dentre 8 biopsiados resultou citogeneticamente normal e foi transferido em ciclo diferido. A paciente teve gravidez e parto a termo, de criança do sexo masculino. Pudemos identificar 38 aneuploidias nos sete blastocistos restantes, determinando a origem parental: oito trissomias maternas, oito paternas e 14 monossomias maternas. Cinco das oito trissomias maternas mostravam má-segregação em alelo paterno 1:1:1 e três mostravam o padrão 2:1. Os mesmos padrões de segregação foram observados nas trissomias de origem paterna. Concluimos que as aneuploidies tiveram contribuição de ambos os membros do casl, em proporção igual. Este desfecho nos leva a refletir nas ovodoações para mulheres acima de 40 anos, quando o companheiro tem idade similar.
Palavras-chave: PGS, biopsia de trofoderma, idade paterna avançada, aborto de repetição, aCGH.
RESUMEN
Es el reporte de un procedimiento PGS realizado por CGH en una pareja de 44 y 46 años que consulta por abortos espontáneos con aneuploidías recurrentes. Solamente un blastocisto de ocho estudiados resultó tener un cariotipo molecular masculino normal, el cual fue transferido y dio lugar a un recien nacido normal con cariotipo convencional normal en sangre de cordón umbilical. En los restantes siete blastocistos aneuploides se intentó determinar el origen de las mismas. Se hallaron 38 aneuploidías. Con la utilización de STRs marcados ligados a los cromosomas involucrados y posterior qf-PCR se logró determinar el origen parental de treinta de aneuploidías, de las cuales fueron: 8 trisomías maternas, 8 paternas y 14 monosomías maternas. Cinco de las ocho trisomías maternas mostraron un patrón alélico de segregación anormal 1:1:1 y tres un patrón 2:1. El mismo patrón de segregación se observó para las trisomías paternas. Por lo tanto, ambos miembros de la pareja contribuyeron con la misma proporción de aneuploidías. Este hallazgo nos obliga a reflexionar cuando se ofrece la alternativa de la ovodonación cuando su pareja es mayor de 40 años.
INTRODUCTION
It is recognized that less than 50% of the fertilization in vivo may lead to baby born. The in vitro fertilization rate is much less; it is between 5% to 30% (Edmonds DK et al., 1982). Several factors are responsible for this low reproductive efficiency, but the most important is the high rate of embryonic chromosome abnormalities of meiotic origin, which varies between 25% and 75% (Hassold T et al., 2007). In women with normal karyotype, the oocyte aneuploidy rate is between 20% and 50% according to the women’s age (Kuliev A and Verlinsky J, 2004; Sandalinas M et al., 2002; Pelliestor F et al., 2003; Fraguoli E et al., 2006; Guitierrez-Mateo C et al., 2004; Hassold T et al., 2007; Sheer G et al., 2007) while in men with normal karyotype and normal spermiogram, aneuploidy is much lower, between 5% and 10% regardless the age (Fonseka KG and Griffin DK, 2011). The lethality of the chromosomal abnormalities is high and most embryos are lost in preimplantional stage. The ones that are not lost at this stage are responsible for chromosomal disorders in the newborn, being 0.67% prevalence (Hassold Tand Hunt P, 2001). Since the aneuploidies and polyploidies affect a large proportion of preimplantation embryos, the application of PGD for the screening of aneuploidies (PGS) should improve pregnancy rate and decrease abortion rate. The PGS was primarily performed by FISH analyzing five chromosomes and it was then extended to reach twelve or more chromosomes (Munné S et al., 2005). Currently there is another technique available; the comparative genomic hybridization performed on a metaphase (m-CGH) or over a platform, which constitutes human chromosome fragments cloned in bacteria (BAC array CGH). This allows the enumeration of all chromosomes and also the detection of small imbalances in each one, with a resolution that might be better than the standard karyotyping. This technique however does not distinguish ploidy, balanced structural rearrangements, or the origin of the aneuploidies.
The combination of the aCGH with quantitative fluorescent PCR allows to partly solve the limitations of the aCGH.
A PGD is performed on a couple with iterative pregnancy loss by recurring aneuploidy using the embryonic molecular karyotyping, complemented with the study of polymorphic chromosome markers or STRs (single tandem repeats) in order to assert and identify the origin of aneuploidy.
PATIENTS AND METHODS
This is a 44 year old female and 46 year old male couple that has been pursuing an ongoing pregnancy for the last six years. They have already completed seven ICSI procedures, as he has an astenozoospermia and bilateral varicocele. She, on the other hand, had no relevant history, presented normal hormonal dosages and normal antral follicle counts. The karyotype couple was normal. Out of the seven ICSIs performed, they achieved pregnancy five times, but ended in clinical abortions between the eighth and eleventh week of gestation. Four of them were cytogenetically studied, and all were diagnosed as aneuploidies: a trisomy 21, a trisomy 15, a trisomy 16 and another with an extra chromosome of the group G. The couple consulted us for a PGS and they went through six ICSIs, but decided not to carry out the PGS due to the quality of the few developed embryos. However two of them achieved pregnancy, but again finished in clinical abortions in the eighth week of pregnancy, one euploid and one with a trisomy 13. The couple decided to keep trying with ICSI procedures, but unlike the previous ones, it was suggested to culture the blastocysts stage and only when they achieved an adequate number, then perform the PGS by aCGH. Two ICSIs were performed and the couple produced four blastocysts in each one on day five. Each blastocyst underwent trophectoderm biopsy. On day three, all embryos with more than five cells received a hole in the pellucid zone with a single seven millisecond laser shot (Octax® 400x lense) to encourage the hatching and the protrusion of the trophectoderm to facilitate biopsy. The cutting and removal of the trophectoderm cells were performed by using two laser shots of 10 milliseconds on both sides of the protruded trophectoderm. Each blastocyst biopsied was then vitrified in a cryotop and the removed cells were washed three times in biopsy medium before being placed in a 2 ml Eppendorf tube with 1 μl of biopsy medium, which was kept in the freezer to make the array CGH with 24 Sure Aneuploidy V3 of BlueGnome ®. The QF-PCR was performed in the amplified DNA from the trophectoderm cells with aneuploid chromosome STRs (# 1, # 2, # 5, # 6, # 7, # 8, # 9, # 10, # 11, # 12, # 13, # 14, # 15, # 16, # 18, # 19, # 20 and # 21). A number of STRs were tested and the ones informative were selected (table 1). The reaction mixture contained a buffer of PCR 5X, dNTPs 10 mM, STRs 0.5 μM and Taq polymerase 5 U/μl. The cycling conditions were 95 °C/5 min. followed by 35 cycles to 95°C/5 min., 55°C/30s, 72°C/30seg. and a final extension to 72°C/5 min. The amplicons obtained were analyzed by capillary electrophoresis in a 310 ABI-Prism and interpreted in accordance with Fig. 1.
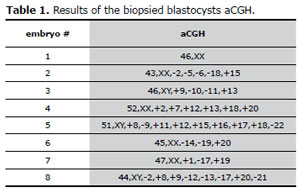
Table 1. Results of the biopsied blastocysts aCGH.
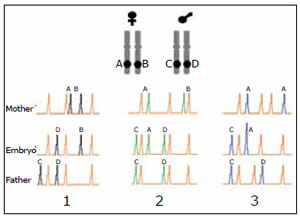
Figure 1. Schematic representation of quantitative fluorescent PCR (QF-PC) using a polimorphic marker (STR) linked to a particular chromosome.
The transfer was made in a subsequent cycle to the stimulation cycle. The endometrium was prepared with estrogens and progesterone. The only normal aCGH blastocyst was warmed and transferred to the uterus after culturing for four hours post thawing (Fig.2). The luteal phase was supplemented with estrogen and progesterone. The pregnancy test was done 10 days post transfer, which was positive, and was corroborated through a vaginal sonogram three weeks later. The woman delivery, at 39 weeks of pregnancy, a normal male child. The conventional karyotype in the blood of the umbilcal cord corroborated the result obtained with the biopsy of trophoectoderm.
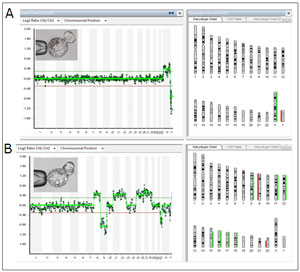
Figure 2. Blastocysts with a normal and an abnormal aCGH(A) The aCGH performed in the removed herniated trophectoderm cells shows a female normal karyotype.(B) The aCGH performed in the removed herniated trophectoderm cells shows nine aneuploides. Seven of them are trisomies of chromosomes 8, 12, 15, 16, 17, 18, and two are monosomies of chromosomes 9 and 22.
RESULTS
Table 1 shows the results of embryonic karyotypes inferred by the aCGH. They all had multiple anomalies with the exception of embryo # 1. The one with the least anomalies had three and the ones with the most had nine aneuploidies (see fig.1). All chromosomes were involved in aneuploidies, with the exception of chromosomes #3, #4 and the sex chromosomes.
Table 2 lists the alleles of the STRs linked to abnormal chromosomes found in the aneuploid blastocysts studied by fluorescent quantitative PCR (QF-PCR). The alleles found revealed the parental aneuploidy origin, and according to the allelic pattern, we could identify in which meiosis phase the error possibly occurred
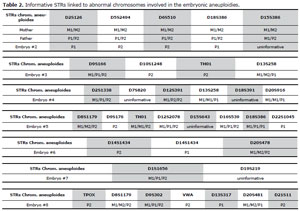
Table 2. Informative STRs linked to abnormal chromosomes involved in the embryonic aneuploidies.
When both members of the couple have different alleles for a given STR, one can identify the parental origin of aneuploidy and furthermore infer in which meiotic anaphase the error occurred, assuming non allelic exchange. In column 1 a normal meiotic division is represented, where each parent contributes a single allele of the STR, the B of the mother and the D of the father (pattern 1:1). Column 2 represents a meiotic error occurred in the first division of the spermatocyte in which the father contributed two alleles of the STR, the C and D, and the mother with the A allele (trisomy of paternal origin that occurred in the first meiotic division, pattern 1: 1: 1). Column 3 represents a meiotic error that occurred in the second division of the oocyte, in which the mother contributed with two equal chromosomes with the A allele (isodisomy) and the father with the chromosome having the allele C (trisomy of maternal origin originated in the second meiotic division of the oocyte, pattern 2: 1).
with the exception of the alleles of the STR of chromosome 15 (embryo # 2), 7-18 (embryo # 4), 15 (embryo # 5) and the 19 in the # 7 embryo, as well as the abnormalities of chromosome 17 (see fig.2).
The origin of the embryonic aneuploidies detected with the aCGH is summarized in table 3. It can be seen that both members of the couple contributed disomies produced by probable errors in both parents during meiosis, but there were no paternal nullisomies.
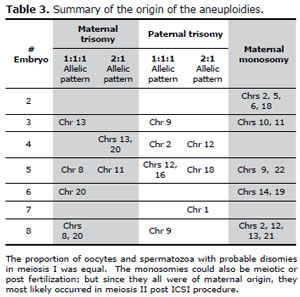
Table 3. Summary of the origin of the aneuploidies.#Embryomaternal trisomy
DISCUSSION
It is known that with the increasing age of women, the ovarian follicular reserve is reduced and/or the response to ovarian stimulation becomes inadequate. The couple, due to their age, had a high chromosome risk during fertilization, mainly because of the oocyte aneuploidies. We can say that the patient, despite her age, still has a good follicular reserve with a good response to ovarian stimulation. The women’s response has not been compromised yet since two cycles of stimulation produced eight blastocysts. However the intrinsic quality of majority of the oocytes was compromised, as well as the spermatozoa that gave origin to the embryos. Out of the eight achieved blastocysts, only one had a normal aCGH. The high rate of aneuploidy found in the present case report has also been communicated by other authors (Alfarawati S et al., 2011). It was widely accepted that the blastocyst aneuploidy rate is much lower than in cleaved embryos. However, the experience gained recently with aCGH in blastocyst biopsy shows that there is a significant rate of aneuploidy, which increases with the age of the woman (Gutierrez-Mateo et al., 2011).
In the present study we have determined the parental origin of the aneuploid chromosomes and we are trying to establish in which meiotic division the error occurred, according to previous communications that used QF-PCR for aneuploidy screening (Katz-Jaffe MG et al., 2004; Diego-Alvarez et al., 2005; Machatkova M, et al., 2005; Diego-Alvarez et al., 2006; Fiorentino et al., 2010; Vahad et al., 2010). We used STRs linked to chromosomes with numerical abnormalities, similar to those used in tests for paternity (see table 4).
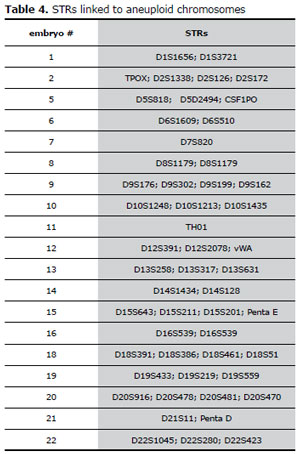
Table 4. STRs linked to aneuploid chromosomesembryo
Without doubt this methodology is a good tool to determine the parental origin of the aneuploidies, but its use to deterrmine if the error occurred in the first or second meiotic division is controversial.
If one assumes that non allelic exchanges occurred between chromatids and the mechanism of malsegregation is the classical non disjunction, one can infer in which division the error occurred. But if the malsegregation is due to a precocious separation of sister chromatids, we can not affirm in which meiotic division the error occurred. If no allelic exchanges occured, the allele pattern 1:1:1 indicates that the trisomy was caused by a malsegregation during anaphase I, by classical non disjunction or by a premature separation of one chromatid (see fig. 3 and 4). On the other hand, if an allele exchange occurred, the pattern 1:1:1 may indicate an error in the first anaphase, by non-disjunction or premature separation of sister chromatid, or by non disjunction during the second meiotic division (see fig 3, 4, 5 and 6). When the trisomy shows an allele pattern 2:1 several mechanisms may explain the pattern: 1) a non disjunction during second meiotic anaphase without alleles interchanged , 2) a non disjunction after fertilization and 3) a precocious separation of sister chromatid with or without alleles interchanged (see fig. 4, 5, 6 and 7). Taking into account the aforementioned limitations, a way to bypass them is through the use of several STRs to determine in which division the error occurred. In the present study, the use of polymorphic markers of aneuploid chromosomes allowed us to identify most of the maternal or paternal origin of aneuploidy as described in Fig.1. In some embryos, it was not possible to identify the aneuploidies of chromosomes 7, 15, 18 and 19 because the results were inconclusive. The study of chromosome 17 was not possible because of the lack of an informative marker.
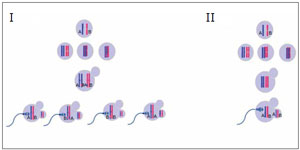
Figure 3. non-disjunction during meiosis I of the oocyte with (I) and without (II) alleles interchangedThe oocyte is disomic (retains the two chromosomes), while the first polar body is nullisomic.To the left (I) is shown an allelic exchange. When the oocyte is penetrated by sperm and resumes the meiosis II, three possibilities of segregation can occur: A: B (or B: A), A: A and B: B, which originate an allelic pattern 1:1:1 or 2:1 in the zygote, respectively. To the right (II) meiosis I nondisjunction without allelic exchange is observed. When the oocyte resumes the meiosis II generates a disomic oocyte (A:B), which leads a 1:1:1 trisomic zygote.
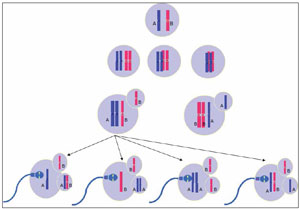
Figure 4. Premature separation of sister chromatids with alleles exchanged during prophase I.The diagram illustrates the four possibilities of segregation during meiosis I and then six possibilities of segregation during the second meiosis for every primary oocyte. Three of the ova correspond to an euploidy rescue, which originate a normal euploid zygotes; two disomic secondary oocytes with an allelic pattern 1: 1, which lead a trisomic zygotes with a 1:1:1 allelic pattern; and finally a disomic ovum, but with a double dose of only one allele, which originates a trisomic egg with a 2:1 allelic pattern.
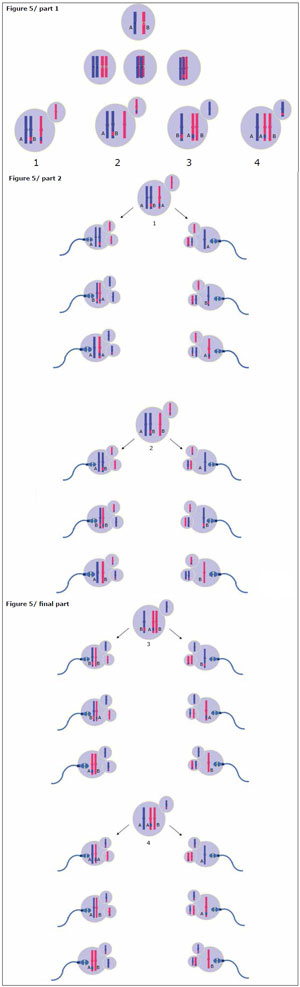
Figure 5. Precocious separation of sister chromatids without alleles exchanged during meiosis IThe diagram illustrates the four possibilities of segregation during meiosis I and then the 24 possibilities of segregation during the second meiosis for every primary oocyte. Twelve ova correspond to an euploidy rescue, which lead a normal euploid zygotes, eight disomic ova with an allelic pattern 1:1, which originate a trisomic zygotes with a 1:1:1 allelic pattern; and four also disomic but with a double dose of only one allele, which create a trisomic zygotes with a 2:1 allelic pattern.
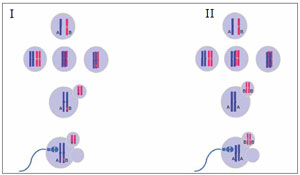
Figure 6. non-disjunction during the second meiotic division of the oocyte with (I) or without (II) alleles interchanged. When the oocyte resumes the meiosis II, the gamete retains the two chromatids (Isodisomic) and the second polar body is nullisomic. On the left side (I), an allelic interchange has occurred (between alleles A and B), so the zygote will present an allelic pattern 1:1:1. On the right side (II), the alleles exchange doesn’t occur and the zygote allelic pattern observed will be 2:1.
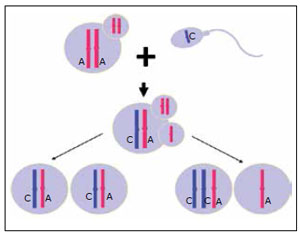
Figure 7. Mitotic Division Post FertilizationColumn 1 shows a normal mitotic division that has a STR allele from each parent (A:C). A non disjunction is represented in column 2, having two chromosomes of the same parent, similar to an error in meiosis II (without alleles exchange), originating an allelic pattern 2:1 (A:A:C).
All aneuploid embryos had several trisomies coexisting with monosomies, except embryo # 2 which did not have any trisomies but instead four monosomies, and embryo # 7 that only showed a trisomy. As you can see in table 3, embryo # 2 presented four (2, 5, 6 and 18) monosomies of maternal origin; embryo # 3 had a maternal trisomy of chromosome 13 with a 1:1:1 allelic pattern, a paternal trisomy 9 with an allelic pattern 1:1:1 and two monosomies (10 and 11) of maternal origin. Embryo # 4 presented a maternal trisomy 13 and 20 with an allelic pattern 2:1, a paternal trisomy 2 with a 1:1:1 pattern and a paternal trisomy 12 with a pattern 2:1. Embryo # 5 showed a maternal trisomy 8 with a 1:1:1 allelic pattern, a maternal trisomy 11 with an allelic pattern 2:1, a paternal trisomy 12 and 16 that presented an allelic pattern 1:1:1, a paternal trisomy 18 with an allelic pattern 2:1 and the monosomies 9 and 22 of maternal origin; embryo # 6 showed a maternal trisomy 20 with an allelic pattern 1:1:1 and two monosomies of maternal origin (chromosomes 14 and 16) ; embryo # 7 showed a trisomy 1 of paternal origin with an allelic pattern 2:1; and embryo # 8 showed maternal trisomies 8 and 20 that presented a 1:1:1 allelic pattern, a paternal trisomy 9 with an allelic pattern 1:1:1, and four monosomies of maternal origin ( chromosomes 2, 12, 13 and 21). The coexistence of trisomies and monosomies could be attributed to the increased susceptibility of the aneuploid zygotes, producing more mitotic errors than normal zygotes or, due to suboptimal conditions of in vitro development, from fertilization to the blastocyst stage or an inadequate competence of the oocyte cytoplasm. While clinical embryologists always overestimate that the IVF laboratory suboptimal conditions are the cause of aneuploidies, we should not forget that most of the chaotic abnomalities are found in dismorphic or arrested embryos. In spite of our study being performed on blastocysts, the majority of the embryos had chaotic chromosomic abnormalities.
Another surprising fact was that all monosomies found were from maternal origin.
Such monosomies could be originated by a premature separation of maternal chromatids (fig.8) as well as the result of post fertilization errors. Since absent chromosomes corresponded to the woman, and in the present case the IVF procedure was an ICSI, we can also speculate that the monosomies could be originated during the second meiotic division of the oocyte as a result of the ICSI “trauma”.
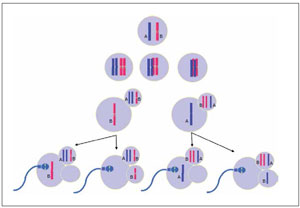
Figure 8. Monosomies originated by Premature separation of sister chromatidsThe diagram illustrates the two possibilities of segregation during meiosis I and then the two possibilities of segregation during the second meiosis for every primary oocyte. Two ova result nullisomic and two euploids.
It is well recognized that most of the autosomic trisomies are from maternal origin, while most of the sex chromosomes anomalies are from paternal origin (Martin RH and Rademaker AW, 1992; Hassold et al., 1996; Freeman et al., 2000). In this case no sex chromosome abnormalities were identified and several autosomic trisomies were found.
The reason for this communication was to show the equal contribution of parental aneuploidies in most blastocyst studied. There are contradictory papers about the increase risk of aneuploidies with the advanced paternal age. The majority of the authors suggest that the rate of aneuploidy in spermatozoa is not related to age but rather to the spermatozoa quality (Wiener-Megnazi Z et al., 2012; Martin RH et al., 1995). The patient at the time of ICSI was 46 years old and the only altered semen parameter was motility, presumably due to bilateral varicocele. Seven out of the eight blastocysts originated by the couple had chaotic aneuploidies (87.5%). In five of seven blastocysts the anomalies were from paternal origin and four of them were probably originated during the meiosis I.
The high percentage of aneuploidies found may be due to; the advanced age of both members of the couple, to a chromosomal instability, to a meiotic mutation in the male which contributes to a higher rate of non-disjunction, to suboptimal conditions of in vitro development or to the ICSI per se that could promote the loss of chromosomes during anaphase II.
It is very difficult to speculate with the experience of a single case, but the significant paternal contribution found in this report should make us consider the single indication of oocyte or sperm donation in couples more than 40 year old.
Acknowledgments:
The authors thank Soledad Aylward and Karen Jury for the translation of the document.