JBRA Assist. Reprod. 2012;16(03):91-95
ARTIGO ORIGINAL
doi: 10.5935/1518-0557.2012.16.3.02
Oocyte dysmorphisms influence chromosomal abnormalities of in vitro-generated embryos
Dismorfismos oocitários influenciam anormalidades cromossômicas em embriões gerados in vitro
aSapientiae Institute - Educational and Research Center in Assisted Reproduction Rua Vieira Maciel, 62 Sao Paulo - SP, Brazil, ZIP: 04503-040
bFertility - Assisted Fertilization Center Av. Brigadeiro Luis Antônio, 4545 Sao Paulo - SP, Brazil, ZIP: 01401-002
ABSTRACT
Objective: To verify the relationship between oocyte dysmorphisms and chromosomal abnormalities of in vitro-derived embryos.
Methods: This retrospective study enrolled 189 intracytoplasmic sperm injection cycles associated with preimplantation genetic aneuploidy screening. The relationship between oocyte dysmorphisms and embryonic chromosomal abnormalities was evaluated. The influence of oocyte dysmorphisms on embryonic chromosomal aneuploidy was assessed using logistic regression and results are expressed as odds ratio (OR), confidence intervals (CI) and p-values.
Results: Increased cytoplasmic granularity (OR: 1.68, CI: 1.10-2.56), enlarged (OR: 1.52, CI: 1.10-2.11) and granular perivitelline space (OR: 1.52, CI: 1.11-2.10) were determinants of increased likelihood of embryonic chromosomal alteration. Considering only female patients aged ≤ 35 years, enlarged perivitelline space (OR: 2.20, CI: 1.11-4.37) was determinant of the likelihood of embryonic chromosomal alteration.
Conclusions: Oocyte cytoplasmic granulation, enlarged and granular perivitelline space are associated with increased probability of aneuploidy in the developing embryo.
keywords: aneuploidy, intracytoplasmic sperm injection, oocyte morphology, embryo, preimplantation genetic screening.
RESUMO
Objetivo: Verificar a relação entre os dismorfismos oocitários e as anormalidades cromossômicas dos embriões gerados in vitro.
Métodos: Este estudo retrospectivo incluiu 189 ciclos de injeção intracitoplasmática de espermatozóides associados ao diagnóstico genético preimplantacional. A relação entre os dismorfismos oocitários e as anormalidades cromossômicas embrionárias foi avaliada. A influência dos dismorfismos oocitários na aneuploidia embrionária foi avaliada por meio de regressão logística e os resultados foram expressos como odds ratio (OR), intervalos de confiança (IC) e valores de p.
Resultados: A granulação citoplasmática (OR: 1,68, IC: 1,10-2,56), o aumento do espaço perivitelíneo (OR: 1,52, IC: 1,10-2,11) e a granulação do espaço perivitelíneo (OR: 1,52, IC: 1.11-2.10) foram determinantes para a maior probabilidade de alteração cromossômica embrionária. Considerando apenas as pacientes com idade ≤ 35 anos, o aumento do espaço perivitelíneo (OR: 2,20, IC: 1,11-4,37) foi determinante da probabilidade de alteração cromossômica embrionária.
Conclusões: A granulação citoplasmática, o aumento e a granulação do espaço perivitelíneo estão associados com uma probabilidade aumentada de aneuploidia no embrião em desenvolvimento.
Palavras-chave: aneuploidia, injeção intracitoplasmática de espermatozóide, morfologia oocitária, embrião, diagnóstico genético preimplantacional.
INTRODUCTION
Oocyte maturation relies on a poorly-understood, complex cascade of events involving the coordination of both nuclear and cytoplasmic changes. Despite the extrusion of the first polar body (IPB) indicates meiosis, it is not sufficient to ensure that the meiotic apparatus of the oocyte has progressed correctly to metaphase II (MII) (De Santis et al. 2005). Disturbance or asynchrony of cytoplasmic and nuclear maturation has been shown to result in different morphologic abnormalities in oocytes (Eichenlaub-Ritter et al. 1995; Hassan-Ali et al. 1998; Loutradis et al. 1999).
With the introduction of intracytoplasmic sperm injection (ICSI) (Palermo et al. 1992), which requires the removal of cumulus cells, it is possible to visualize new information regarding oocyte morphology. Good quality human MII oocytes should have a clear, moderately granular cytoplasm that does not contain inclusions, a small perivitelline space (PVS) containing a single unfragmented IPB and a round, clear, colorless zona pellucida (ZP) (Veeck 1988). Oocyte dysmorphisms can be observed in more than half of collected oocytes (Ebner et al. 2006). nonetheless, the importance of oocyte morphology on in vitro fertilization (IVF) is still a matter of debate. Several studies have reported on the relationship, or lack thereof, between oocyte morphology and ICSI outcomes (De Sutter et al. 1996; Verlinsky and Kuliev 1996; Serhal et al. 1997; Balaban et al. 1998; Loutradis et al. 1999; Kahraman et al. 2000; Meriano et al. 2001; Otsuki et al. 2004; De Santis et al. 2005; Yakin et al. 2007). Furthermore, an association between oocyte dysmorphisms and embryonic aneuploidy has been proposed (Van Blerkom and Henry 1988). nonetheless, despite many studies investigating this possible association (Van Blerkom and Henry 1992; Alikani et al. 1995; Verlinsky and Kuliev 1996; Kahraman et al. 2000; Balaban and Urman 2003; Otsuki et al. 2004; Yakin et al. 2007), the issue remains unclear.
Therefore, this study was designed to verify the possible relationship between oocyte dysmorphisms and embryonic chromosomal abnormalities in the developing embryo.
MATERIALS AND METhODS
Experimental design, patients and inclusion criteria We retrospectively identified 189 cycles, performed from January 2007 to December 2009, which fulfilled the following inclusion criteria: ICSI followed by preimplantation genetic aneuploidy screening (PGS). To minimize the influence of male factor infertility, all cases of sperm concentration less than 1 x 106 M/mL and sperm motility less than 20% were excluded from the study.
In the first analysis, the relationship between oocyte intracytoplasmic dysmorphisms (increased cytoplasmic granularity, vacuoles in the ooplasm, and aggregates of smooth endoplasmic reticulum clusters (SER) in the ooplasm) and extracytoplasmic dysmorphisms (fragmented IPB, PVS granularity and enlarged PVS), and embryonic chromosomal alteration was evaluated in the 189 identified cycles (834 oocytes/ embryos). In order to exclude the influence of maternal age on embryonic chromosomal abnormalities, a second analysis of the above-described protocol was performed including only the embryos from female patients up to 35 years old (n=57, 326 oocytes/embryos).
A written informed consent was obtained, in which patients agreed to share the outcomes of their own cycles for research purposes, and the study was approved by the local institutional review board.
Controlled ovarian stimulation
Controlled ovarian stimulation was achieved by long-term pituitary down-regulation using a gonadotropin-releasing hormone agonist (GnRH agonist, Lupron KitTM, Abbott S.A Societé Française des Laboratoires, Paris, France). This procedure was followed by ovarian stimulation with recombinant follicle-stimulating hormone (FSH) (Gonal-F®, Serono, Geneve, Switzerland). Oocyte retrieval was performed 35 hours after the administration of recombinant human chorionic gonadotrophin (rhCG, OvidrelTM, Serono, Geneve, Switzerland), through transvaginal ultrasonography.
Preparation of oocytes
Retrieved oocytes were maintained in human tubal culture medium (HTF, Irvine Scientific, Santa Ana, USA) supplemented with 10% Human Synthetic Albumin (HSA, Irvine Scientific, Santa Ana, USA) and covered with mineral oil (OvoilTM, Vitrolife) for 2-3 hours before cumulus cell removal. Surrounding cumulus cells were removed after exposure to a HEPES buffered medium containing hyaluronidase (80IU/mL, Irvine Scientific, Santa Ana, USA). The remaining cumulus cells were then mechanically removed by gently pipetting with a hand-drawn Pasteur pipette (Humagen Fertility Diagnostics, Charlottesville, USA).Oocyte morphology was assessed using an inverted nikon Diaphot microscope (Eclipse TE 300; nikon®, Tokyo, Japan) with a Hoffmann modulation contrast system under 400X magnification, just before sperm injection (3-4 hours after retrieval). The following oocyte dysmorphisms were recorded: (i) increased cytoplasmic granularity, (ii) vacuoles in the ooplasm, (iii) smooth endoplasmic reticulum clusters (SER) in the ooplasm (iv) large PVS size, (v) PVS granularity and (vi) fragmented IPB. Oocytes that were observed to have released the first polar body were considered mature and were used for ICSI.
Intracytoplasmic sperm injection, fertilization and embryo development
Oocytes were transferred into a micro-injection dish, prepared with 4 µL droplets of buffered medium (HEPES, Irvine Scientific, Santa Ana, USA), and covered with mineral oil on a heated stage at 37.0 ± 0.5°C of an inverted microscope. Approximately 16 hours after ICSI, fertilization was confirmed by the presence of two pronuclei and the extrusion of the second polar body. Embryos were kept in a 50ul drop of HTF medium supplemented with 10% HSA covered with mineral oil in a humidified atmosphere under 6% CO2 at 37ºC until transfer. Embryonic development was further evaluated on days 2, 3, 4 and 5.
Embryo biopsy
Embryos reaching at least the 5-cell stage on day 3 of development were biopsied by laser zona drilling using a 1.48µm Infrared Diode Laser (Octax Laser Shot System, MTG, Bruckberg, Germany) and returned to culture. Only one blastomere was removed per embryo. The definition of a successful biopsy was the removal of a cell without lysis, so that the cell could be used for fixation and analysis. If the blastomere was not successfully biopsied or fixed, a second blastomere was taken, but only if the embryo initially had ≥6 blastomeres.
Blastomere fixation and fluorescent in situ hybridization (FISh)
The blastomere of an embryo was fixed on a slide according to the HCI/Tween 20 method, as previously described (Dumoulin et al. 2005). A two-round FISH procedure was performed, which allowed for the detection of chromosomes X, Y, 13, 18 and 21 (Multivision PGT Probe Panel; Vysis, Downers Grove, IL, USA) in the first round and chromosomes 16 and 22 in the second round. The hybridization solution for the second round was prepared by mixing a probe for chromosome 16 (Vysis, Satellite II DnA/D16Z3 probe, Spectrum Orange) and a probe for chromosome 22 (Vysis, LSI 22, 22q11.2, Spectrum Green). The FISH procedure was performed as previously described (Van de Velde et al. 2000). The results of the first and second round were analyzed using a fluorescence microscope.
FISh scoring criteria
At diagnosis, embryos were considered normal when two gonosomes and two chromosomes 13, 16, 18, 21 and 22 were present. Several FISH procedure-related factors were responsible for unreliable results secondary to the following mechanisms: damaged or incomplete nuclei, nuclei covered with debris or nuclei without clear signals. This was referred to as FISH failure.
Embryo transfer
Embryo transfer was performed on day 5 by using a soft catheter with transabdominal ultrasound guidance. One to three embryos were transferred per patient.
Clinical Follow-up
A pregnancy test was performed 12 days after embryo transfer. All women with a positive test had a transvaginal ultrasound scan 2 weeks after the positive test; a clinical pregnancy was diagnosed when the fetal heartbeat was detected. Pregnancy rates were calculated per transfer. Miscarriage was defined as spontaneous abortion before 20 weeks of gestation.
Statistical analysis
The influence of oocyte dysmorphisms on embryonic chromosomal aneuploidy was assessed using a binary logistic regression analysis, and results are expressed as odds ratio (OR), confidence intervals (CI) and p-values. Results were considered to be significant at the 5% critical level (p< 0.05). Data analysis was carried out using MInITAB Software.
RESULTS
Patients 26-45 years old (n=189)
General characteristics of the first analysis are shown in Table I. A total of 892 embryos were biopsied. The removal of an extra blastomere was necessary in 18 (2.0%) embryos. Due to technical issues, including hybridization failure, signal overlapping yielding false-negative results and split or diffuse signals, the embryo biopsy failed in 58 (6.5%) cells. Out of 834 embryos with FISH results, 441 were aneuploid (52.9%).
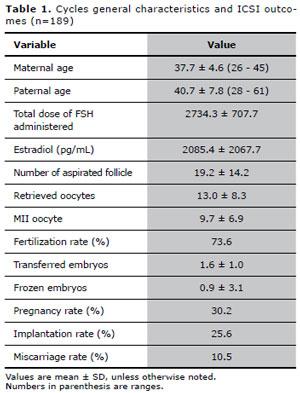
Table 1. Cycles general characteristics and ICSI outcomes (n=189)
Considering only the oocytes which originated embryos with FISH results, increased cytoplasmic granularity was observed in 690 (82.7%) oocytes, vacuoles in 24 (2.9%), aggregates of SER in 13 (1.6%), fragmented IPB in 227 (27.2%), PVS granularity in 445 (53.4%) and enlarged PVS in 347 (41.6%) oocytes.For this group of patients, our results showed that oocytes displaying cytoplasmic granularity, PVS granularity or enlarged PVS were associated with a significantly increased chance of generating a chromosomally abnormal embryo. The influence of oocyte dysmorphisms on embryonic chromosomal alteration, evaluated through binary logistic regression, is shown in Table II. It was not possible to calculate the effects of SER, due to a low number of interactions in the binary logistic regression.
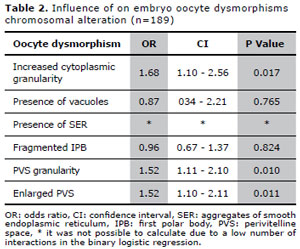
Table 2. Influence of on embryo oocyte dysmorphisms chromosomal alteration (n=189)
Patients aged ≤ 35 years old (n=57)
General characteristics of the second analysis are show in Table III. A total of 348 embryos were biopsied. The removal of an extra blastomere was necessary in 8 (2.3%) embryos. Due to technical issues, including hybridization failure, signal overlapping yielding false-negative results and split or diffuse signals, the embryo biopsy failed in 22 (6.3%) cells. Out of 326 embryos with FISH results, 145 were aneuploid (44.5%).Considering only the oocytes that originated embryos with FISH results, increased cytoplasmic granularity was observed in 178 (54.6%) oocytes, vacuoles in 9 (2.8%), aggregates of SER in 9 (2.8%), fragmented IPB in 63 (19.3%), PVS granularity in 114 (35.0%) and enlarged PVS in 68 (20.8%) oocytes.For this group of patients, our results showed that the oocytes presenting with enlarged PVS was associated with a significantly increased chance of generating a chromosomally abnormal embryo. The influence of oocyte dysmorphisms on embryonic chromosomal alteration, evaluated through binary logistic regression, is shown in Table IV. It was not possible to calculate the effects of vacuoles and SER, due to a low number of interactions in the binary logistic regression.
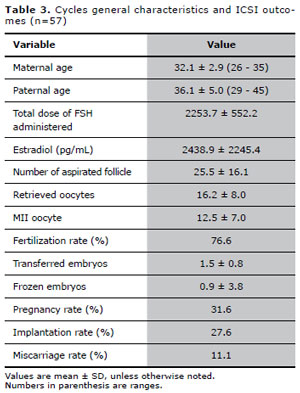
Table 3. Cycles general characteristics and ICSI outcomes (n=57)
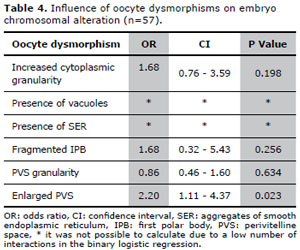
Table 4. Influence of oocyte dysmorphisms on embryo chromosomal alteration (n=57).
DISCUSSION
In this study we showed that oocyte dysmorphisms have an adverse effect on embryonic chromosomal status, regardless of maternal age. Oocytes presenting cytoplasmic granularity, PVS granularity or enlarged PVS were associated with an increased chance of chromosomally abnormal embryo generation in female patients aged 26-45 years old. To exclude the effects of advanced maternal age, the oocytes and derived embryos from a subgroup of female patients aged ≤ 35 years were analyzed. The association between enlarged PVS and embryonic chromosomal alteration was maintained in this subgroup of patients.
Increased cytoplasmic granulation of an oocyte may be a sign of cytoplasmic immaturity (Kahraman et al. 2000); nevertheless, it is not known what factors are responsible for this kind of oocyte dysmorphism. Ovarian stimulation for IVF allows a more synchronous cohort of follicles to be recruited, but some oocytes may come from slower developing follicles. These oocytes’ cytoplasms would be at different maturation stages when exposed to hCG for the resumption of meiosis (Xia 1997). Therefore, nuclear and cytoplasmic maturation can proceed one without the other, resulting in compromised embryos (Eppig et al. 2002).
It is not clear how cytoplasmic abnormalities compromise zygotic or embryonic development. During the completion of meiosis I and the transition to the metaphase II stage, synchronous cytoplasmic maturation, mitochondrial changes, protein synthesis and cytoskeletal changes take place (Scott 2000). Because the proteins responsible for sperm chromosome decondensation, release of protamines, DnA repair, chromosome remodeling and assembly of organelles are accumulated in the cytoplasm (Collas 1998), the quality of the cytoplasm plays a pivotal role in male pronuclear assembly and the fertilization process (Banerjee et al. 2000). It has been suggested that chromosomal abnormality may be one reason for cytoplasmic granulation (Kahraman et al. 2000). The effects of cytoplasmic granularity on embryo chromosomal status have been previously investigated. Yakin et al. (2007) showed that despite a 20% higher aneuploidy rate was found among embryos derived from these oocytes, the difference did not reach statistical significance. A specific kind of cytoplasmic granularity, known as centrally located granular cytoplasm (CLCG), can be observed in rare situations. Kahraman et al. (2000) reported that 53% of the embryos derived from the group of oocytes with CLGC were aneuploid.
In addition, defects in the cytoplasmic machinery may block gene activation. Wells et al. (2005) assessed a small number of embryos derived from centrally granulated oocytes for gene expression on Day 3 and found that this type of oocyte morphology is associated with altered gene expression.
The fate of the oocytes with extracytoplasmic abnormalities is still unknown. It was demonstrated that the aneuploidy rate for the group of embryos derived from oocytes with cytoplasmic abnormalities was higher compared with either the group of embryos derived from oocytes with extracytoplasmic abnormalities or the group of embryos derived from oocytes with normal morphology (Balaban and Urman 2003). nonetheless, Rienzi et al. (2008) showed that not only cytoplasmic abnormalities but also extracytoplasmic abnormalities are indicators of oocyte competence, reflecting a decreased potential of fertilization and compromised pronuclear morphology.
In this study we observed that the presence of an enlarged PVS was a determinant of embryonic chromosomal alteration regardless of maternal age. The PVS is known to start forming when meiotic maturation resumes and to enlarge in matured oocytes at the metaphase II stage (Kaufman et al. 1989). It has been suggested that an enlarged PVS may be related to an overmaturity of the oocyte (Mikkelsen and Lindenberg 2001; Rienzi et al. 2008), which, in turn, causes polar body retention and influences the incidence of embryonic aneuploidy (Badenas et al. 1989).
Granularity of the PVS, in this study, also influenced the incidence of embryonic chromosomal alteration. However, this association was not seen in younger women. There is evidence that some PVS inclusions are the remnants of coronal cell processes (York et al. 1993; Sathananthan 1997). Figueira et al. (2009) observed that the total dose of follicle stimulating hormone (FSH) was associated with the presence of PVS granularity. It is well known that poor ovarian response to stimulation protocols increases as women get older (Veleva et al. 2005); therefore, those patients are usually stimulated with high doses of gonadotrophins (Kumbak et al. 2008). Because we did not observe an association between PVS granularity and embryonic chromosomal alteration in younger patients, it could be suggested that high doses of gonadotrophin administered in the older patients resulted in the maturation of oocytes with increased PVS granularity and probability of embryonic chromosomal alteration in the developing embryo.
In spite of a good experimental design, our study possesses three other drawbacks, as follows:
(i) We did not perform polar body chromosomal analysis; therefore, we cannot state for certain that embryonic aneuploidy was related to the oocyte. The exclusion of patients with severe male factors may be minimized but not totally refused the impact of spermatozoa-related embryonic aneuploidy.
(ii) We performed single blastomere biopsy for embryonic chromosomal analysis. However, because there is no convincing data demonstrating the safety of double over single-blastomere biopsy (Van de Velde et al. 2000; Edwards and Hansis 2005), we perform single blastomere biopsy in our center.
(iii) Due to a small number of interactions between the oocytes displaying aggregates of SER and embryo chromosomal status in the first analysis and between the oocytes presenting aggregates of SER or vacuoles in the second analysis, it was not possible to verify the relationships between these two important dysmorphisms and embryonic chromosomal alteration.
CONCLUSION
In conclusion, we showed that oocyte cytoplasmic granulation, enlarged PVS and PVS granularity are associated with an increased probability of aneuploidy in the developing embryo. Therefore, the practical and inexpensive practice of oocyte morphological evaluation should not be discontinued.
Balaban B and Urman B. Embryo culture as a diagnostic tool. Reprod Biomed Online. 2003;7:671-82.
Collas P. Cytoplasmic control of nuclear assembly. Reprod Fertil Dev. 1998;10:581-92.
Sathananthan AH. Ultrastructure of the human egg. Hum Cell. 1997;10:21-38.
Scott LA. Oocyte and embryo polarity. Semin Reprod Med. 2000;18:171-83.
Veeck LL. Oocyte assessment and biological performance. Ann n Y Acad Sci. 1988;541:259-74.